Across Africa, the virtual abandonment of handheld large cutting tools such as handaxes, and an increased emphasis on prepared core technologies and hafting,
marked a profound technological reconfiguration of hominin material
culture. These technological changes, which define the transition to the
Middle Stone Age (MSA), seem to have occurred across
Africa at a broadly similar time; for example at ∼300 ka both at Jebel
Irhoud, where they are found with early H. sapiens fossils [16xThe age of the hominin fossils from Jebel Irhoud, Morocco, and the origins of the Middle Stone Age. Richter, D. et al. Nature. 2017;
546: 293–296
Crossref | PubMed | Scopus (47) | Google ScholarSee all References][16], and at Olorgesailie in East Africa [30xLong-distance stone transport and pigment use in the earliest Middle Stone Age. Brooks, A. et al. Science. 2018;
360: 90–94
Crossref | PubMed | Scopus (4) | Google ScholarSee all References][30], and at ∼280 ka in southern Africa at Florisbad [31xDirect dating of the Florisbad hominid. Grün, R. et al. Nature. 1996;
382: 500–501
Crossref | PubMed | Scopus (113) | Google ScholarSee all References][31]. Currently, the earliest dates in West Africa are younger, at ∼180 ka, but the region remains very poorly characterized [32xThe West African Stone Age. Scerri, E.M.L.
Google ScholarSee all References][32]. The MSA is associated with H. sapiens fossils, but both H. naledi and H. heidelbergensis probably persisted into the late Middle Pleistocene.
Clear
regionally distinctive material culture styles, typically involving
complex stone tools, first emerged within the MSA. For example, the
Central African MSA includes heavy-duty axes, bifacial lanceolates,
backed flakes and blades, picks and segments, probably from at least the
late Middle Pleistocene [33xAcross rainforests and woodlands: a systematic reappraisal of the Lupemban Middle Stone Age in Central Africa. Taylor, N. : 273–300
Crossref | Scopus (7) | Google ScholarSee all References][33]. In the Late Pleistocene,
grassland and savannah expansion in North Africa led to dense human
occupation associated with specific regional technological features such
as tanged implements (Figure 2Figure 2) [34xThe North African Middle Stone Age and its place in recent human evolution. Scerri, E.M.L. Evol. Anthropol. 2017;
26: 119–135
Crossref | PubMed | Scopus (2) | Google ScholarSee all References][34].
At approximately the same time there was an emergence of comparably
distinctive industries in parts of southern Africa. As in North Africa,
some of these industries are also associated with other aspects of
complex material culture such as ochre, bone tools, shell beads, and
abstract engravings (Figure 2Figure 2) [35xIdentifying
early modern human ecological niche expansions and associated cultural
dynamics in the South African Middle Stone Age. d’Errico, F. et al. PNAS. 2017;
114: 7869–7876
Crossref | Scopus (9) | Google ScholarSee all References][35].

Figure 2
Middle
Stone Age Cultural Artefacts. (A–D) Bifacial foliates from northern
Africa (A, Mugharet el Aliya; B–D, Adrar Bous). (E–G) Bifacial foliates
from southern Africa (Blombos Cave). (H,I) Tanged tools from northern
Africa. (J) Segmented piece bearing mastic residue from southern Africa
(Sibudu). (K) Engraved ochre fragment (Blombos Cave). (L–N) Engraved
ostrich eggshell fragments from southern Africa (Diepkloof). (O,P) Bone
points from southern Africa (Sibudu and Blombos Cave, respectively). (Q)
Bone point from northern Africa (El Mnasra). (R–V) Perforated Trivia gibbosula
shells from northern Africa (R,S, Grotte de Pigeons; T–V, Rhafas, Ifri
n’Ammar, and Oued Djebbana, respectively). (W–Aa) Perforated Nassarius kraussianus shells from Blombos Cave. (Ab) Conus ebraeus
shell bead (Conus 2) from southern Africa (Border Cave). (Ac) Ochre
fragment shaped by grinding from southern Africa (Blombos Cave). All
scales are 1 cm. Boxed items indicate rescaled artefacts. Images
reproduced, with permission, from (A–D, H, I) The Stone Age Institute;
(E–G, J–P, Ac) from [35xIdentifying
early modern human ecological niche expansions and associated cultural
dynamics in the South African Middle Stone Age. d’Errico, F. et al. PNAS. 2017;
114: 7869–7876
Crossref | Scopus (9) | Google ScholarSee all References][35]; (Q) from [47xOsseous projectile weaponry from early to late Middle Stone Age Africa. Backwell, L. and d’Errico, F. : 15–29
Crossref | Scopus (8) | Google ScholarSee all References][47]; and (R–Ab) from [35xIdentifying
early modern human ecological niche expansions and associated cultural
dynamics in the South African Middle Stone Age. d’Errico, F. et al. PNAS. 2017;
114: 7869–7876
Crossref | Scopus (9) | Google ScholarSee all References, 47xOsseous projectile weaponry from early to late Middle Stone Age Africa. Backwell, L. and d’Errico, F. : 15–29
Crossref | Scopus (8) | Google ScholarSee all References, 48xAdditional evidence on the use of personal ornaments in the Middle Paleolithic of North Africa. d’Errico, F. et al. PNAS. 2009;
106: 16051–16056
Crossref | PubMed | Scopus (140) | Google ScholarSee all References].
Such
regionalization is typically linked with the emergence of ‘modern’
cognition. However, it arguably also reflects the interaction between
demographic variables (e.g., increased population density) [36xLate Pleistocene demography and the appearance of modern human behavior. Powell, A. et al. Science. 2009;
324: 1298–1310
Crossref | PubMed | Scopus (438) | Google ScholarSee all References, 37xThe Still Bay and Howiesons Poort: ‘Palaeolithic’ techno-traditions in southern Africa. Henshilwood, C.S. J. World Prehist. 2012;
25: 205–237
Crossref | Scopus (54) | Google ScholarSee all References, 38xCoalescence and fragmentation in the late Pleistocene archaeology of southernmost Africa. Mackay, A. et al. J. Hum. Evol. 2014;
72: 26–51
Crossref | PubMed | Scopus (61) | Google ScholarSee all References] and the learned traditions of long-lived regional subpopulations or demes (Figure 2Figure 2).
For example, northern and southern Africa, apart from being
geographically distant, were also separated by environmental factors as a
consequence of the expansion and contraction of forests in equatorial
Africa, synchronous with amelioration in northern Africa. Other factors,
such as habitat variability and adaptation to local environmental
conditions, are also likely to play some role in material culture
diversification.
Although geographical differences are
clear at the continental scale, localized spatial patterning is harder
to discern. Similarities between regions may have been produced by
occasional contact or by convergent adaptation to common environmental
conditions. In East Africa, for example, although there is certainly
some variation, there appears to be underlying continuity in material
culture throughout much of the MSA (e.g., [39xStone tool assemblages and models for the dispersal of Homo sapiens out of Africa. Groucutt, H.S. et al. Quat. Int. 2015;
382: 8–30
Crossref | Scopus (28) | Google ScholarSee all References][39]). In many regions, ‘generic’ MSA assemblages that do not carry an obvious signal of regionalization are common [40xEcological
risk, demography, and technological complexity in the Late Pleistocene
of Northern Malawi: implications for geographical patterning in the
Middle Stone Age. Thompson, J. et al. J. Quat. Sci. 2017;
33: 261–284
Crossref | Scopus (1) | Google ScholarSee all References][40].
In a cognitive model, these differences suggest that not all these
early populations manifested a ‘modern mind’. However, such assemblages
are augmented by shifting frequencies of tool types that appear to be
spatially or temporally indicative, and likely reflect demographic
factors. In some parts of Africa, the full suite of generalized MSA
characteristics continues largely unchanged until the
Pleistocene/Holocene boundary [41xPersistence
of Middle Stone Age technology to the Pleistocene/Holocene transition
supports a complex hominin evolutionary scenario in West Africa. Scerri, E.M.L. et al. J. Archaeol. Sci. Rep. 2017;
11: 639–646
Google ScholarSee all References][41],
matching the morphological patterns, and suggesting that the end of the
MSA may have been as structured and mosaic-like as its beginnings. This
view has support from LSA material culture. Despite superficial
similarities in LSA lithic miniaturization, the cultural record shows
continued differentiation and derivation into the Holocene, supporting
the biological evidence for variable population dynamics that did not
result in wide-scale homogenization [42xA demographic perspective on the Middle to Later Stone Age transition from Nasera rockshelter, Tanzania. Tryon, C.A. and Faith, J.T. Philos. Trans. R. Soc. 2016;
B 371
Crossref | PubMed | Scopus (9) | Google ScholarSee all References][42].
The
reasons for, and therefore implications of, the geographic and temporal
structuring of MSA cultural diversity are still poorly characterized
and likely reflect several processes. These include adaptations to
different environments [43xThe evolution of the diversity of cultures. Foley, R.A. and Mirazón-Lahr, M. Philos. Trans. R. Soc. 2011;
366: 1080–1089
Crossref | PubMed | Scopus (44) | Google ScholarSee all References][43]. Long-term, large-scale population separation may also have been the norm for much of Pleistocene Africa (Box 1;
i.e., isolation by distance and isolation by habitat, representing null
models to be rejected). Rare and spatially explicit models exploring
Pleistocene technological innovations have also linked cultural
complexity with variation in regional patterns of population growth,
mobility, and connectedness (e.g., [36xLate Pleistocene demography and the appearance of modern human behavior. Powell, A. et al. Science. 2009;
324: 1298–1310
Crossref | PubMed | Scopus (438) | Google ScholarSee all References, 44xCulture, population structure, and low genetic diversity in Pleistocene hominins. Premo, L.S. and Hublin, J.-J. PNAS. 2009;
106: 33–37
Crossref | PubMed | Scopus (62) | Google ScholarSee all References, 45xPopulation density, mobility, and cultural transmission. Grove, M. J. Archaeol. Sci. 2016;
74: 75–84
Crossref | Scopus (4) | Google ScholarSee all References]), supported by evidence of long-distance transfer of stone raw material (e.g., [46xThe
earliest long-distance obsidian transport: evidence from the ∼200 ka
Middle Stone Age Sibilo School Road Site, Baringo, Kenya. Blegen, N. J. Hum. Evol. 2016;
103: 1–19
Crossref | PubMed | Scopus (7) | Google ScholarSee all References][46]).
Box 1
+
Box 1
Isolation by Distance (IBD)
IBD
is the expectation that genetic differences correlate positively
geographic distances as a consequence of the fact that mating is more
likely to occur at shorter than longer distances. This concept, although
well established in genetics, is rarely applied to Pleistocene
archaeological and human fossil material, despite its potential value as
a null model for observed cultural or morphological differences between
materials from different sites [49xEstimating mobility using sparse data: application to human genetic variation. Loog, L. et al. PNAS. 2017;
114: 12213–12218
Crossref | PubMed | Scopus (0) | Google ScholarSee all References][49].
Because
archaeological problems are concerned with identifying the processes
that generate observed patterns of cultural variation over time and
space, the lack of equivalent null models is particularly problematic.
Processes generating cultural variation constitute a complex balance
between patterns of inherited knowledge, local innovation, modes of
cultural transmission, local adaptation, and shifting population
dynamics (e.g., population size, density, or mobility [36xLate Pleistocene demography and the appearance of modern human behavior. Powell, A. et al. Science. 2009;
324: 1298–1310
Crossref | PubMed | Scopus (438) | Google ScholarSee all References][36]).
Without null models of ‘cultural similarity’ as a baseline, it is
difficult to escape simplistic, narrative inferences about the past.
Similarly, human fossil data can be interpreted in several ways
depending on the taxonomy employed, but spatial variation is likely to
relate to the same factors that influence genetic similarities between
regional populations [50xDistance from Africa, not climate, explains within-population phenotypic diversity in humans. Betti, L. et al. Proc. R. Soc. 2009;
B 276: 809–814
Crossref | Scopus (77) | Google ScholarSee all References][50].
In
an archeological context, the expectation of an IBD model is that
cultural similarity will decrease with distance, with a degree of
spatial autocorrelation (e.g., [51xIsolation-by-distance, homophily and ‘core’ vs. ‘package’ cultural evolution models in Neolithic Europe. Shennan, S.J. et al. Evol. Hum. Behav. 2015;
36: 103–109
Abstract | Full Text | Full Text PDF | Scopus (16) | Google ScholarSee all References][51]).
This expectation represents the simplest explanation of the observed
variation. If this null model does not provide an adequate explanation
of the data, more complex models can be invoked to explain patterns
observed either in the residuals from the null model or in the raw data
as a whole. For example, a more complex population structure can be
theoretically differentiated from an IBD model if patterns of spatial
autocorrelation are discrete rather than continuous (Figure IFigure I),
pointing towards the formation of distinct biological or cultural
clusters which may correlate with other features (e.g., genetic,
morphological, or environmental). Many factors could promote the
formation of such clusters, including assortativity by cultural
similarity, or conformity to local norms of, for example, tool
production.
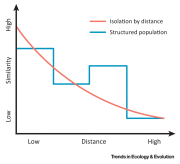
Figure I
Simple
IBD Model with Cultural Data. Note that similarity can increase with
distance under some circumstances, for example when similar habitats are
separated by considerable distances, with areas of different habitat
types being located between them.
Major
new archaeological research directions should include: (i) unraveling
the relative contributions of different African regions/habitats to
recent human evolution; (ii) understanding shifting patterns of
population structure through the differential appearance, expansion,
contraction, and disappearance of regionally distinct artefact forms (Box 1);
and (iii) exploiting the growing interface between archaeology,
ecology, morphology, and genetics to explore the extent to which
material culture patterning is coupled or decoupled from these
associated (but potentially independent) axes.



 Article Info
Article Info










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}